
Por: Priscila Silva Grijó Farani
- Os ovos de Schistosoma mansoni
O primeiro estudo sobre a estrutura do ovo de S. mansoni, revelou que a camada logo abaixo da casca do ovo maduro (camada de Reynold) apresenta microfibras inseridas em uma matriz granular. Abaixo desta, cercando completamente o miracídio e outras estruturas está presente um fino envelope (envelope de von Lichtenberg) de origem celular escamosa, que parece envolvido na regulação do transporte dos antígenos do ovo e outros produtos embrionários para fora e nutrientes para dentro do ambiente onde o miracídio está localizado1,2. A formação do granuloma, principal causa de morbidade da doença, é estritamente dependente da viabilidade dos ovos maduros, não ocorrendo ao redor dos ovos imaturos3.
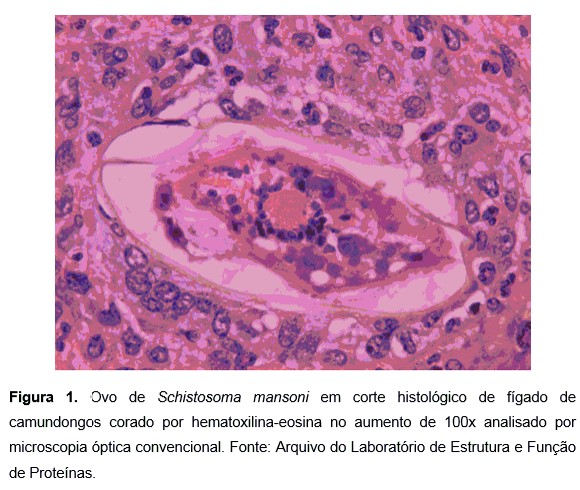
A casca do ovo consiste em três camadas: camada externa microespinhosa, camada densa intermediária medial e camada densa interna. A camada chamada de Reynold (Reynold`s layer), desenvolve-se subjacentemente à casca do ovo e é compreendido de ramificações de filamentos bem compactos. Uma camada única de células escamosas, também chamada de envelope de von Lichtenberg está intimamente aderido à camada de Reynold1. Estas suas estruturas apresentam similaridades ontogênicas, estruturais e funcionais ao longo do desenvolvimento do miracídio3. A camada citoplasmática compreendida entre o fluido extracelular do hospedeiro e o miracídio em desenvolvimento forma uma barreira contra a difusão passiva simples, sugerindo que macromoléculas complexas como antígenos esquistossomóticos, sofrem transporte ativo ou até mesmo seletivo para dentro e para fora do ovo1.
O ovo de S. mansoni recém-depositado é relativamente pequeno, com aproximadamente 110 µm, possuindo um conteúdo altamente granulado e sem sinais visíveis do complexo arranjo de membranas e vesículas que cercam o miracídio no ovo maduro. O conteúdo do ovo imaturo compreende células vitelinas nucleadas com citoplasma rico em mitocôndrias, gotículas lipídicas, vesículas de diversos tipos de inclusões granulares, possivelmente para armazenamento de glicogênio2.
Com o desenvolvimento do ovo, placas dispersas de citoplasma se tornam uma camada contínua separando as células vitelinas da casca e delimitando completamente os conteúdos do ovo. Neste estágio intermediário, a camada apresenta uma superfície lisa interna e externa, o citoplasma granular contém pequenas mitocôndrias e estruturas membranosas. Uma pequena quantidade de material floculento está presente no espaço entre a camada e a casca2.
No ovo completamente desenvolvido, o envelope possui um citoplasma altamente diferenciado em que o núcleo já pode ser identificado confirmando dessa forma a natureza celular da camada. O citoplasma é dividido em regiões distintas, em que largas vesículas estão presentes, provavelmente com o intuito de armazenar lipídios, enquanto a maior área é ocupada por micro agregados de material granular. Outra característica marcante é a expansão do reticulo endoplasmático rugoso, principalmente no envelope, que é indicativo de atividade de síntese proteica. O envelope de von Lichtenberg encontra-se completamente desenvolvido entre a casca e a camada de Reynold, apresentando grandes quantidades de material granular. Os microsespinhos da superfície externa casca circundam a camada de material e o miracídio fica adjacente ao envelope com alguns de cílios em contato direto, sendo banhado em um ambiente de material denso e livre2.
Estágios de embriogênese no ovo de S. mansoni
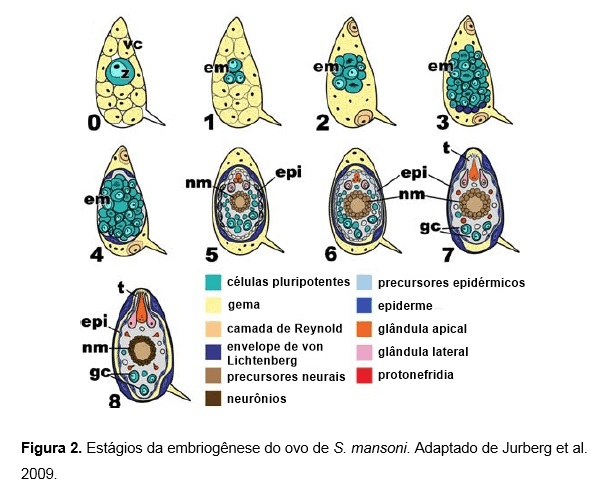
Apenas uma pequena parte do ciclo de desenvolvimento do ovo ocorre dentro da fêmea do verme adulto e dois estágios podem ser definidos: o estágio pré-zigótico e o estágio zigótico. No estágio pré-zigótico os oócitos deixam o ovário projetando seu citoplasma na forma de pseudópodes. No estágio zigótico, também denominado estágio 0, o zigoto torna-se cercado por células vitelinas, que são menores que o zigoto propriamente dito e localizam-se centralmente apresentando núcleo eosinofílico. O zigoto, juntamente com as células vitelinas são empurradas pela fêmea por meio de contrações musculares para o oótipo onde a casca do ovo é formada, e o ovo passa pelo útero e é liberado através do gonóporo para o ambiente externo, no entanto ainda não se apresenta completamente formado3.
Estágio 1
Os ovos recém-colocados são carregados passivamente pela corrente sanguínea até que fiquem aprisionados em pequenas veias. Neste estágio o embrião é muito pequeno e isso dificulta sua visualização. A primeira divisão produz dois blastômeros pouco diferentes em tamanho, exibindo um núcleo grande e redondo com um nucléolo proeminente e intensamente basófilo3.
O citoplasma exibe características de intensa atividade de síntese protéica, e as células vitelinas exibem inicialmente um citoplasma vacuolado, mas que começam a se fundir em um único sincício conforme a evolução do ovo3. Ao final deste estágio o embrião apresenta uma forma discoide se tornando levemente visível na área central do ovo, ocupando cerca de metade de seu comprimento transversal3.
Estágio 2
O embrião inicial cresce em tamanho, mas as células embrionárias e o sincício permanecem morfologicamente semelhantes ao estágio anterior3.
As células vitelinas fundem-se completamente e alguns macrômeros parecem soltar-se do pólo do embrião aderindo-se a casca do ovo, dando origem à camada de Reynold. Estes macrômeros possuem núcleo grande e redondo, com nucléolo redondo, intensamente basófilo e proeminente, com citoplasma escasso e basófilo. Ao final deste estágio, uma massa sólida denominada estereoblástula ocupa todo o comprimento transversal do ovo3.
Estágio 3
O embrião agora apresenta-se como uma massa celular clara e desenvolve-se em tamanho, assumindo uma forma alongada e ocupando cerca de dois terços do comprimento longitudinal do ovo. O embrião ainda apresenta certa dificuldade de visualização e é possível identificar o início da diferenciação celular, com a presença de pelo menos dois tipos celulares distintos3.
As células do tipo I apresentam uma morfologia imatura, enquanto as células do tipo II (identificadas como possivelmente sendo neuroblastos) concentram-se no centro do embrião, apresentando um núcleo pequeno e irregular, citoplasma escasso e nucléolo puntiforme3.
As células embrionárias da periferia do ovo apresentam-se comprometidas com a diferenciação do envelope de von Lichtenberg e seus núcleos apresentam-se ligeiramente achatados3.
Estágio 4
Neste estágio a diferenciação do envelope de von Lichtenberg se torna aparente com a formação do sincício separado ao redor do ovo e apresenta uma distribuição dispersa de ribossomos. O núcleo permanece com característica achatada e um nucléolo lateral pronunciado. A camada de Reynold funde-se com o sincício e desaparece progressivamente durante o desenvolvimento3.
Os núcleos vitelinos são visíveis nos pólos do embrião e a massa celular central surge de um grupo celular de forma arredondada com núcleo pequeno, heterocromático e nucléolo puntiforme (células do tipo II) 3.
Estágio 5
Neste estágio o embrião ocupa quase toda a área interna do ovo, apesar de alguns restos vitelinos ainda estarem presentes nos pólos. Os ovos apresentam crescimento progressivo e a camada de Reynold assume uma característica fina, granular e anucleada, dificultando cada vez mais sua visualização3.
A massa celular neural aumenta de tamanho expandindo dessa forma sua região central e acelular. A epiderme começa a diferenciar-se das outras células primordiais do embrião, que fundem-se umas com as outras no intuito formar o sincício localizado logo abaixo do envelope de von Lichtenberg. Essas células primordiais apresentam citoplasma altamente basófilo, indicativo de síntese proteica. Na região anterior do embrião é possível visualizar duas células imaturas e largas, que aparecem em cada lado da massa celular neural3.
Estágio 6
Neste estágio as células imaturas localizadas na porção lateral anterior do ovo exibem citoplasma granulado e já podem ser identificadas como precursoras das glândulas laterais do miracídio. O núcleo permanece grande e redondo, com nucléolo proeminente mas sem delimitação de heterocromatina3.
Outro grupo identificado de células imaturas darão origem a glândula apical da larva madura, enquanto a epiderme da região anterior do embrião projeta-se discretamente para frente, definindo uma estrutura denominada terebratório3.
Na região medial-posterior do embrião aparecem células imaturas que correspondem às células germinativas do miracídio. O envelope de von Lichtenberg apresenta as mesmas características do estágio anterior e já não é mais possível identificar a camada de Reynold3.
Ao final desta etapa, o epitélio epidérmico se torna claramente diferenciável do mesênquima e os precursores musculares começam a diferenciar-se abaixo da epiderme3.
Estágio 7
O embrião alongado ocupa todo o volume do ovo, mas ainda não é possível observar nenhum movimento. Durante este estágio a massa neural sofre condensação nuclear, dando origem a um pequeno núcleo grosseiramente redondo e alongado3.
É possível observar um aumento no volume do neurópilo e na porção medial anterior do embrião, as células germinativas não apresentam diferenças em relação ao estágio anterior. A casca do ovo mostra-se mais espessa dos pólos, e já é possível observar a musculatura subepidermal do miracídio3.
Estágio 8
O embrião ocupa todo o interior do ovo e é possível observar intensa motilidade do miracídio, com destaque para as contrações musculares e os batimentos ciliares. O miracídio encontra-se pronto para nascer, se encontrar-se em condições favoráveis3.
Uma ou duas camadas de neurônios encontram-se organizadas perifericamente ao redor do neurópilo. O terebratório, a glândula apical e a as glândulas laterais estão completamente formadas na região anterior do miracídio e é possível observar evidências de secreções glandulares no interior do ovo3.
A superfície epidérmica é formada por placas epidermais ciliadas, que produzem um fino sincício que possui núcleo redondo e basófilo. Nenhuma célula mitótica está presente no embrião, e as células germinativas apresentam-se em grupo de quatro ou cinco células no pólo medial-posterior. A camada de Reynold não apresenta diferenças em suas características, mas o envelope de von Lichtenberg apresenta-se discretamente mais fino3.
– O granuloma de Schistosoma mansoni
O granuloma esquistossomótico (Figuras 3 e 4) é uma estrutura organoide, dinâmica e híbrida, formada por componentes pertencentes tanto ao ovo quanto ao hospedeiro, possuindo uma fase de maturação e involução 4,6. A formação do granuloma ocorre partindo de uma fase inicial desordenada e, pouco a pouco, forma-se um esqueleto randômico entremeado com células que se intensifica gradualmente, apresentando um arranjo semelhante a uma malha de forma concêntrica ou radial, definindo áreas claras dentro dos granulomas 5,6.
A estrutura do granuloma caracteriza-se pela presença de grupos de células pertencentes ao sistema fagocitário mononuclear, podendo ou não haver a presença de outros tipos celulares 6,7. Essas células, influenciadas por forças químicas e biomecânicas, arranjam-se em padrões específicos incluindo a ordenação de diferentes tipos celulares em relação a outras. Os efeitos de agregação e ordenação criam diferentes zonas na estrutura do granuloma, em que a combinação de diferentes tipos celulares formam três áreas distintas: periovular, paracentral e periférica 5.
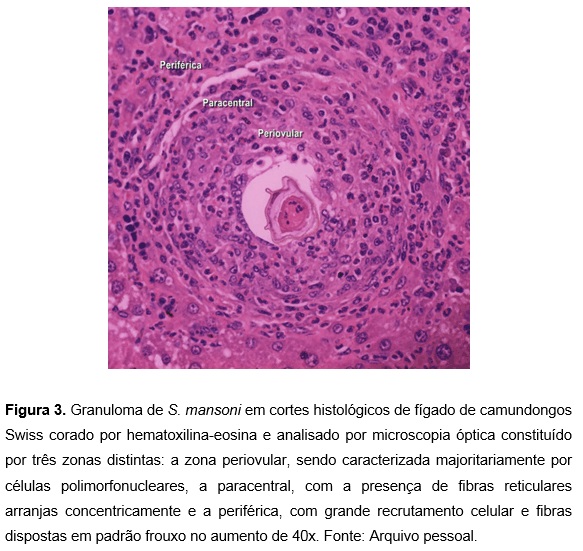
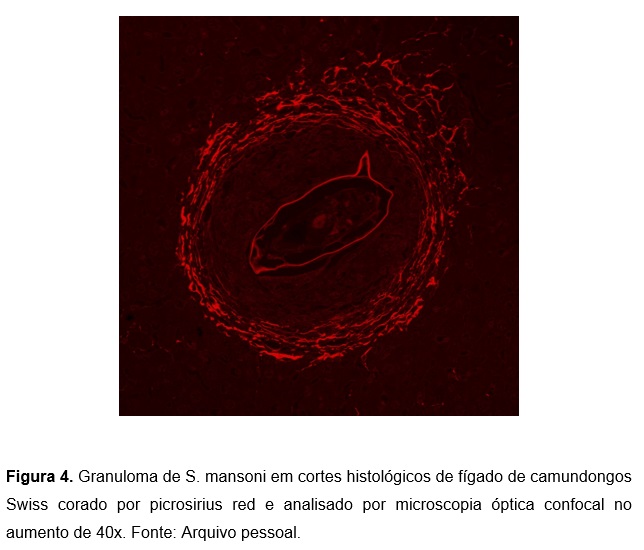
De acordo com Lenzi et al.6, o granuloma de Schistosoma mansoni divide-se nos seguintes estágios:
Estágios pré-granulomatosos
(I) Fracamente/inicialmente reativo: Apresenta acúmulo gradual de células mononucleares, neutrófilos e eosinófilos ao redor do ovo recém-depositado.
(II) Exsudativo: Este estágio promove destruição focal das paredes vasculares envolvidas e do parênquima adjacente, com ocorrência de histólise e microtrombose principalmente no fígado, que se beneficia da “capacidade metastática” dos ovos de S. mansoni em penetrar os vasos sanguíneos e permite que eles se depositem nos tecidos justavasculares, favorecendo também a liberação de ovos nas fezes.
Estágios granulomatosos
(I) Exsudativo-produtivo: Apresenta aparecimento de histiócitos e células epitelióides que substituem gradualmente a zona leucocitária. Fibrócitos também começam a aparecer na periferia da lesão e forma uma zona externa ao redor dos histiócitos e células epitelióides. É caracterizada pela vasta presença de células migrantes (componente exsudativo) e locais que produzem diversos componentes da matriz extracelular (componente produtivo).
(II) Produtivo: O ovo de Schistosoma se torna degenerado e desintegrado, fibrócitos e fibras colágenas se tornam mais proeminentes. Linfócitos, histiócitos, células plasmáticas e alguns eosinófilos formam uma zona adicional na periferia da lesão. Os fibrócitos e as fibras colágenas eventualmente se tornam predominantes nas características do granuloma, enquanto os outros tipos celulares diminuem em número.
(III) Involutivo: Os granulomas apresentam-se bem reduzidos em tamanho e exibem hialuronização das fibras colágenas, enquanto o ovo se encontra geralmente desintegrado. A fase involutiva pode apresentar granulomas com:
dissociação das fibras colágenas
camada espessa de colágeno
camada delgada de colágeno
macrófagos pigmentados
Processo de formação do granuloma
Durante os primeiros dias, o ovo é aprisionado em uma veia aproximadamente do seu tamanho. Então, várias células como monócitos e eosinófilos, com presença de poucos linfócitos, formam um coxim celular ao redor do ovo, chamado também de processo de encapsulação6.
Algumas células formam “pontes” do ovo ao endotélio, e penetram então através da parede do vaso criando uma pequena bainha perivenular e, posteriormente, o número de células infiltradas aumenta e se difunde intensamente para o tecido perivascular, sendo quimioatraídas, juntamente com as células, para os sinusóides do fígado ou outros vasos adjacentes, formando um conjunto frouxo de várias células exsudativas com dimensões que podem atingir cinco vezes o tamanho do ovo6.
Este agregado de células exsudativas ocupa o espaço formado pela histólise focal. Em seguida, a camada medial é formada, rica em fibroblastos e miofibroblastos provenientes da parede do vaso original, e novas fibras colágenas. A construção do emaranhado de fibras da camada medial começa usualmente da parte mais interna da parede vascular residual, na interface entre a cama interna e o começo da cama medial, procedendo em direção à camada mais externa6.
No caso de ovos aprisionados no espaço portal, as fibras são produzidas sobre uma vasta área, rapidamente e ao mesmo tempo. Sendo assim, o processo de produção de fibras pode começar mais cedo juntamente com o processo de encapsulação. Em alguns casos, a construção ocorre ao redor do ovo e o desenvolvimento é para o exterior, simultaneamente, fibras dispersas são geradas na cama externa6.
Em um estágio posterior da fase produtiva-exsudativa, o emaranhado de fibras da camada medial se torna mais compacta, paralela e concêntrica, adquirindo um arranjo circunferencial. Após a morte espontânea do miracídio, a camada interna encolhe, enquanto a camada medial é empurrada para dentro devido à pressão dos tecidos subjacentes. O processo inflamatório desaparece, mantendo apenas alguns poucos macrófagos pigmentados, e a degradação da matriz extracelular começa a predominar sobre a formação da mesma, tendendo a reestabelecer uma relação normal ou quase normal entre estroma/parênquima. Ao final da fase de desintegração, o granuloma desaparece completamente6.
Referências
| 1 Neill P, Smith JH, Doughty BL, Kemp. The ultrastructure of the Schistosoma mansoni egg. J Trop Med Hyg. 1988; 39, 52-65. | |
| 2 Ashton PD, Harrop R, Shah B, Wilson RA. The schistosome egg: development and secretions. Parasitology. 2001; 122, 329-338. | |
| 3 Jurberg AD, Gonçalves T, Costa TA, de Mattos AC, Pascarelli BM, de Manso PP, Ribeiro-Alves M, Pelajo-Machado M, Peralta JM, Coelho PM, Lenzi HL. The embryonic development of Schistosoma mansoni eggs: proposal for a new staging system. Dev Genes Evol. 2009; 219:219-234. | |
| 4 Lins RAB, Cavalcanti CBL, Araújo-Filho JLS, Melo-Júnior MR, Chaves MEC. Distribution of eosinophils at different stages of hepatic granuloma evolution in mice infected with Schistosoma mansoni. Rev Soc Bras Med Trop. 2008 Mar-Apr; 41(2):173-178. | |
| 5 Carvalho OS, Coelho PMZ, Lenzi HL. Schistosoma mansoni & Esquistossomose, uma visão multidisciplinar. Rio de Janeiro: Editora Fiocruz; 2008. | |
| 6 Lenzi HL, Kimmel E, Schechtman H, Pelajo-Machado M, Romanha WS, Pacheco RG, Mariano M, Lenzi JA. Histoarchitecture of schistosomal granuloma development and involution: morphogenetic and biomechanical approaches. Mem Inst Oswaldo Cruz. 1998; 93(1):141-51. | |
| 7 Lenzi HL, Kimmel E, Schechtman H, Pelajo-Machado M, Vale BS, Panasco MS, Lenzi JA. Collagen arrangement in hepatic granuloma in mice infected with Schistosoma mansoni: dependence on fiber radiation centers. Braz J Med Biol Res. 1999 May; 32(5):639-43. | |